

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)
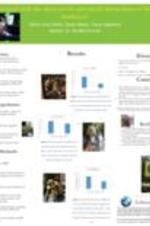
Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)
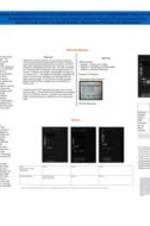
Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)

Enhancing Global Research and Education in STEM at Spelman College (G-STEM)
